
If there is one concept that sits at the heart of almost everything I spent my PhD years thinking about, it is this: proteins fold, and sometimes they fold wrong, and when that happens in the brain, the consequences can be devastating.
Protein misfolding is the unifying mechanism behind some of the most common and most devastating neurodegenerative diseases we know, Alzheimer’s disease, Parkinson’s disease, Huntington’s disease, ALS, and others. Understanding it is not just intellectually fascinating (although it absolutely is, I promise). It is one of the most important frontiers in biomedical research, and I think everyone deserves a real explanation of what is actually happening, not just the vague “protein clumps cause brain damage” summary that usually makes it into mainstream science coverage.
So let’s actually get into it.
Proteins: A Quick Foundation
Before we talk about misfolding, we need to talk about folding, what it is, why it happens, and why it matters so much.
A protein begins as a linear chain of amino acids, assembled by ribosomes in your cells according to instructions encoded in your DNA. That chain is called a polypeptide. But a linear chain of amino acids is not yet a functional protein. To do its job, whether that’s catalysing a chemical reaction, providing structural support, signalling between cells, or any of the thousands of other tasks proteins perform, the chain has to fold into a precise three-dimensional shape.
This folding is not random. It is driven by the physical and chemical properties of the amino acids themselves, particularly their attraction to or repulsion from water, and the various bonds that can form between them. The final folded shape of a protein is generally the lowest-energy conformation available to it, which is why proteins tend to fold consistently and reproducibly into the same shape. That shape is everything. A protein that is folded correctly does its job. A protein that folds incorrectly, or unfolds under stress, cannot.
Helper proteins called chaperones assist with folding and can sometimes refold a misfolded protein back to its correct shape. Your cells also have quality control systems, including the proteasome, which degrades misfolded proteins, and autophagy, which I have talked about before, specifically dedicated to catching and clearing proteins that aren’t right. Under normal circumstances, these systems keep misfolding under control.
In neurodegeneration, they fail to keep up.
What Is Protein Misfolding and Why Does It Happen?
Misfolding occurs when a protein adopts an incorrect three-dimensional conformation, a shape that is stable enough to persist but wrong enough to be non-functional or, worse, actively harmful.
There are several reasons this can happen. Genetic mutations can alter the amino acid sequence in ways that destabilise the normal fold. Oxidative stress, the accumulation of reactive oxygen species that damage cellular components, can chemically modify proteins in ways that disrupt their structure. Simply aging reduces the efficiency of the cellular quality control machinery that normally catches and clears misfolded proteins before they accumulate. And some proteins are just intrinsically prone to misfolding under certain conditions.
What makes misfolded proteins particularly dangerous in the context of neurodegeneration is what happens next: they aggregate.
The Aggregation Problem
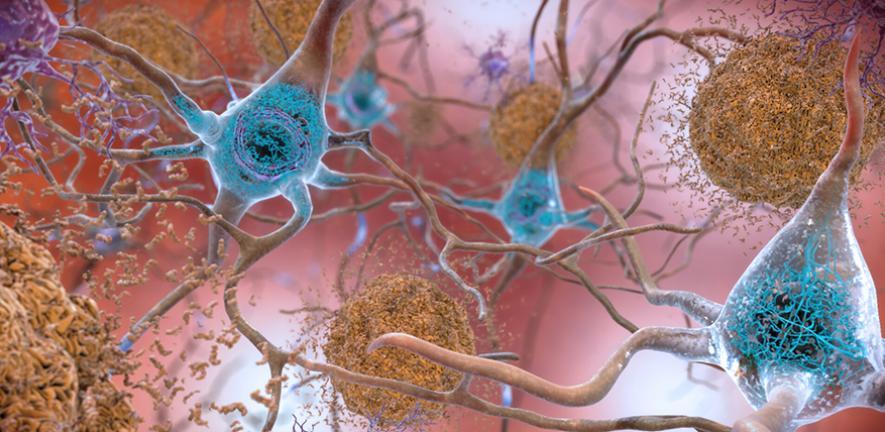
When a protein misfolds, it often exposes hydrophobic regions, parts of the molecule that are normally tucked away in the interior of the correctly folded structure, away from the water-based environment of the cell, because they are repelled by water. Once these hydrophobic patches are exposed, the misfolded protein has a strong tendency to stick to other misfolded proteins that are exposing similar patches. One misfolded protein becomes two stuck together. Two becomes four. The aggregate grows.
These aggregates can take various forms. Some are relatively small and soluble, oligomers, containing just a handful of proteins stuck together. Others grow into larger, more structured deposits called amyloid fibrils, which have a characteristic cross-beta sheet structure: long, ordered fibers made of beta-strand elements from multiple proteins stacking against each other in a ladder-like arrangement. This beta-sheet structure is extraordinarily stable, amyloid fibrils are resistant to most of the cellular machinery that would normally degrade them, and it is a common end state for many different aggregating proteins.
The protein involved differs between diseases. In Alzheimer’s disease, the primary culprits are amyloid-beta peptides (which form the plaques described in the classic disease pathology) and tau protein (which forms the neurofibrillary tangles found inside neurons). In Parkinson’s disease and Lewy body dementia, it is alpha-synuclein that misfolds and aggregates into structures called Lewy bodies. In Huntington’s disease, it is the mutant huntingtin protein. In ALS and frontotemporal dementia, TDP-43 and FUS are among the proteins involved.
Different proteins, different diseases, but the same fundamental story: misfolding, aggregation, cellular dysfunction, and death of neurons.
Why Neurons Are Particularly Vulnerable
You might reasonably wonder: don’t all cells have to deal with misfolded proteins? Why are neurons the ones that suffer so catastrophically in these diseases?
Several reasons, and they compound each other in unfortunate ways.
Neurons are post-mitotic, which means they do not divide. Most cells in your body can dilute the burden of misfolded protein aggregates simply by dividing: when a cell splits, the aggregates are distributed between daughter cells and effectively halved. Neurons cannot do this. Whatever accumulates in a neuron stays there for the life of that neuron, which, in the central nervous system, is intended to be the entire life of the organism.
Neurons are also extraordinarily metabolically demanding. They consume large amounts of energy and produce large amounts of reactive oxygen species as a byproduct. This creates a high-oxidative-stress environment that is particularly hard on proteins, increasing the likelihood of modification and misfolding. At the same time, neurons are large and structurally complex cells with long axons that can extend enormous distances, which places particular demands on the transport and quality control systems that need to function properly throughout the whole cell.
And neurons depend on each other. When one neuron in a circuit dies, it places stress on the others it was connected to. Neurodegeneration tends to spread through connected brain regions, which is why these diseases are progressive.
The Prion-Like Spreading Hypothesis
One of the most significant, and frankly startling, developments in neuroscience research over the last two decades has been the growing evidence that misfolded protein aggregates can spread between cells in a manner that resembles, mechanistically, the behaviour of prions.
Prions are famously the cause of diseases like Creutzfeldt-Jakob disease and bovine spongiform encephalopathy (mad cow disease), and their defining characteristic is that a misfolded protein can act as a template, converting normally folded proteins it contacts into the same misfolded conformation. One misfolded protein, in other words, can seed the misfolding of others around it.
Research now suggests that alpha-synuclein, tau, and amyloid-beta aggregates can behave similarly, that aggregates released from one cell can be taken up by a neighbouring cell and seed new aggregation there. This would explain the characteristic spreading patterns of neurodegenerative diseases through the brain, which tend to follow anatomically connected pathways rather than appearing randomly.
This does not mean Alzheimer’s or Parkinson’s is infectious in the way a cold is infectious. The mechanism is similar but the epidemiology is completely different. However, it has profound implications for thinking about how these diseases progress and how they might be interrupted.
Why This Matters for Treatment
If protein misfolding and aggregation are the root of the problem, then the logical therapeutic strategies are to prevent misfolding, prevent aggregation, promote clearance of aggregates, or interrupt the spread of aggregates between cells. And in fact, most current and emerging therapeutic approaches in neurodegeneration are targeting one or more of these steps.
Immunotherapy approaches, using antibodies to target amyloid-beta or tau aggregates, have been the subject of enormous investment and, recently, some cautious clinical progress. Strategies to boost autophagy or proteasomal clearance are being explored. Small molecules that stabilise the correctly folded form of a protein or prevent the nucleation step of aggregation are active areas of research.
Progress has been slow and humbling, because the brain is the most complex organ we have and because these diseases are often diagnosed long after the damage has already accumulated for years or decades. But the mechanistic understanding is deep and growing, and I remain genuinely optimistic about where the field is heading.
The fact that so many seemingly different diseases converge on the same fundamental mechanism, misfolding, aggregation, cell death, is both sobering and clarifying. It means that understanding one of these diseases deeply enough can illuminate all of them. And that feels worth the effort.







